Как хищники стали травоядными – Наука – Коммерсантъ
Генетические изменения в процессе эволюции животных происходили намного раньше, чем внешние: в ходе крупного международного проекта расшифровки геномов группа российских и зарубежных ученых изобрела метод определения конкретных участков хромосом (или горячих точек), где происходили кардинальные перестройки геномов.
Фото: Юрий Мартьянов, Коммерсантъ / купить фото
Долгое время науке было неизвестно, как и почему за миллионы лет хищные животные становились травоядными копытными, как это произошло, например, с коровой и другими жвачными. Палеонтологи построили свою теорию эволюции, основываясь на постепенных морфологических изменениях, которые подтверждались останками скелетов древних животных. Однако с появлением генетических методов выяснилось, что животные, внешне похожие на ежей или землероек, кардинально отличались между собой по типу питания и образу жизни, поскольку уже тогда они были далекими предками разных современных животных — китов, дельфинов, жирафов, слонов и коров.
Но каким образом это стало возможным?
— Способность переваривать растительную пищу стала величайшим достижением эволюции,— объясняет один из авторов публикации, вышедшей в марте 2019 года в ведущем генетическом научном журнале Genome Research Александр Графодатский из Института молекулярной и клеточной биологии СО РАН.
— У современной коровы в желудке целых четыре отдела, в части которых живут бактерии, которые помогают ей переваривать траву.
Хищники не способны ее переварить. Среди копытных их осталось немного. У кабарги, например, до сих пор сохранились способность есть животную пищу и массивные клыки. Один из древних предков копытных — крошечный зверь под названием оленек — до сих пор существует и проживает в южноазиатских странах, где питается крабами и мелкими грызунами.
В работе, поддержанной проектом РНФ № 16–14–10009, с помощью биоинформатики мы подбирали наборы BAC-клонов из районов, предположительно перестроенных, и которые мы не видели традиционными методами молекулярной цитогенетики. В ИМКБ СО РАН мы локализовали эти клоны на хромосомах видов парнокопытных и смотрели, какие именно «скрытые» перестройки происходили в эволюции. Партнеры из Китая пересеквенировали и анализировали эти районы и выявили многие ранее скрытые геномные события в эволюции китопарноколпытных. Самым интересным стало обнаружение возникающего при этих перестройках своего рода «волнения» геномов, существенным образом изменяющего регуляцию целых групп генов. Вполне вероятно,— и тех, что меняли функцию при изменении типа питания у жвачных. Это предположение окончательно подтвердится после детального и полного изучения именно этой геномно-эволюционной загадки.
За последние годы в отделе разнообразия и эволюции геномов ИМКБ СО РАН собрана коллекция клеток и ДНК более сотни важнейших с точки зрения эволюции видов нового таксона китопарнокопытных, включающего свиней и верблюдов, жирафа и окапи, оленей, антилоп, овец и коров, бегемотов и их родственников — китов и дельфинов. Хромосомы некоторых видов были отсортированы и использованы в качестве молекулярных проб для локализации на хромосомах других видов. Например, хромосома одной коровы метится определенным веществом и локализуется на хромосомах верблюда или кита. Этот метод, называемый хромосомной живописью (или ZooFISH), позволяет определять гомологичные районы хромосом, то есть участки геномов, идентичные по генному содержанию, и их перестройки. Кроме хромосом копытных ученые использовали в качестве зондов и хромосомы человека, что позволило сразу сравнивать гомологичные районы хромосом копытных и наиболее изученного генома человека. Таким образом им удалось определить сходство и различия геномов человека, свиньи, верблюдов, коровы, жирафы и окапи, косули и вилорога, бегемотов и китов, редчайших малазийских оленьков и даже саолы — «тянитолкая» из джунглей Вьетнама, единственный известный науке экземпляр которого есть в российской коллекции.
Хромосомы некоторых видов были отсортированы и использованы в качестве молекулярных проб для локализации на хромосомах других видов. Например, хромосома одной коровы метится определенным веществом и локализуется на хромосомах верблюда или кита. Этот метод, называемый хромосомной живописью (или ZooFISH), позволяет определять гомологичные районы хромосом, то есть участки геномов, идентичные по генному содержанию, и их перестройки. Кроме хромосом копытных ученые использовали в качестве зондов и хромосомы человека, что позволило сразу сравнивать гомологичные районы хромосом копытных и наиболее изученного генома человека. Таким образом им удалось определить сходство и различия геномов человека, свиньи, верблюдов, коровы, жирафы и окапи, косули и вилорога, бегемотов и китов, редчайших малазийских оленьков и даже саолы — «тянитолкая» из джунглей Вьетнама, единственный известный науке экземпляр которого есть в российской коллекции.
Мария Роговая
Про хищников и корону-кровопийцу: моё новое письмо Эволюции: mari_batinak — LiveJournal
Сегодня утром, прочитав одну из последних статей Эволюции, я написала ей очередное письмо.
Мои вопросы касались «хищников» — так Эволюция называет разных любовных аферистов, которые разводят своих жертв на большие деньги.
Вечером она уже опубликовала разбор.
«Глупый ник у автора и глупые вопросы, — написала она обо мне. — Вообще, что касается вопросов про корону, многие читатели отчаянно тупят, потому что очень не хотят с короной расставаться».
Вот само письмо.
Почему у хищников нет короны?
Здравствуйте, Эволюция!
Прочитала ваш недавний пост про корону-кровопийцу, и у меня сразу возник вопрос, который у меня каждый раз появляется, когда я читаю у вас про эту корону.
А как же хищники?
(У хищников нет корон, никаких. Исключение — очень мелкие хищники. У тех иногда мелькает что-то похожее на корону, но конечно не такая корона как у рапанов)
Ведь они сознательно вызывают у людей аддикцию — болезненную привязанность к себе, но при этом у хищников, как я понимаю, нет никакой короны-кровопийцы, иначе, в короне, у них бы ничего не получалось.
(Хищник ничем не отличается от обычного человека и его цель: вызвать симпатию у того, кто ему нравится. Каждый человек хотел бы влюбить в себя того, с кем он желает отношений. У хищника нет цели вызвать именно «болезненную» привязанность. Более того, цель хищника сделать привязанность партнера как можно более приятной для него. Чем счастливей человек и чем больше он рад любить хищника, тем хищнику лучше. Понятно же почему, друзья? Только рапаны могут хотеть, чтобы их партнер испытывал боль. Понимаете почему?)
Выходит, жертвы хищников по-настоящему страдают по ним, когда те отдаляются, ревнуют, боятся потерять и т. д. — то есть переживают весь тот набор чувств, о котором мечтают другие люди в коронах-кровопийцах, чтобы их партнёры чувствовали по отношению к ним. И при этом хищник добивается всего этого от своих жертв сознательно, специально.
(Хищник не добивается боли и страданий. Хищник добивается кайфа и удовольствия партнера. Страх потери — побочный эффект, которого многие хищники, самые хитрые хищники, как раз избегают. Зачем избегают, поясните, друзья? И обратите внимание, какая большая корона-кровопийца у автора письма. Она уверена, что любовь без боли существовать не может)
Зачем избегают, поясните, друзья? И обратите внимание, какая большая корона-кровопийца у автора письма. Она уверена, что любовь без боли существовать не может)
Чем тогда хищник, сознательно вызывающий аддикцию у своей жертвы, и все вышеперечисленные переживания у неё, отличается от человека в короне-кровопийце, который хочет от своих партнёров того же?
(У хищника цель — вызвать кайф. А рапаны вроде автора хотят выносить мозг и мучить людей, поэтому от них сбегают быстро или даже сразу шарахаются. Только увидят перекошенное кислое лицо и торчащие из всех карманов щипцы, и обходят стороной)
Ведь у хищника не должно быть никакой короны?
Почему у хищника всё получается, а у людей в коронах-кровопийцах страдания их партнёров по ним бывают только в их собственных иллюзиях?
И как хищник, осознанно вызывая аддикцию у своего партнёра, умудряется не надевать при этом корону-кровопийцу? Почему у хищников такой короны нет?
(Я другой вопрос задам. Как можно быть настолько бестолковым как автор? Как можно перепутать кайф и боль и считать, что это одно и то же, что люди будут бегать за теми, кто хочет причинять им боль? Как думаете, друзья?)
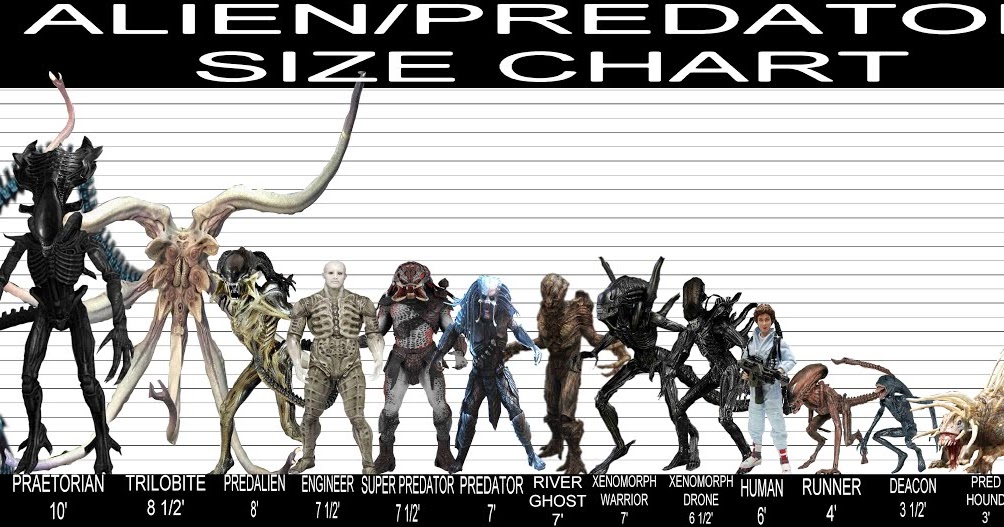
Каждая версия Хищника на протяжении всей истории
Каждая версия Хищника на протяжении всей истории — GameSpotАвтор Дэн Оти по
«Хищник» почти готов, и обзоры готовы. ужасных смертей в истории Хищника и узнайте больше о недавнем споре о Хищнике.
Хищник, несомненно, один из самых знаковых монстров в современном кинематографе. Впервые представленная в классическом Predator 1987 года, эта безжалостная, очень умная машина для убийств прокладывала себе путь через хорошо вооруженный отряд солдат, прежде чем встретиться со столь же культовым Арнольдом Шварценеггером. Успех фильма привел к появлению пяти сиквелов, в том числе грядущего «Хищник », а также обширной вселенной комиксов, романов и видеоигр.
Но, несмотря на блестящий первоначальный дизайн Хищника, первый фильм начал сниматься с совершенно другим героем-инопланетянином. У оригинального костюма Хищника была длинная шея, странная птичья голова и согнутые назад ноги, но технические проблемы с костюмом привели к закрытию производства на несколько месяцев, в то время как Стэн Уинстон, визионер по эффектам макияжа, работал над новым образом, который теперь часть истории кино.
С тех пор внешний вид Хищника менялся от фильма к фильму, и в каждой новой части серии адаптировался и обновлялся базовый дизайн. Некоторые из этих изменений касаются доспехов и вооружения, а иногда и внешнего вида самого пришельца. Часто изменения заключаются в том, чтобы Хищник выглядел страшнее, чем в предыдущем фильме, но иногда требуется более симпатичный Хищник, например, в первых 9.0009 Чужой против Хищника . Сценаристы, художники и дизайнеры комиксов и игр также развернули дизайн в некоторых интересных новых направлениях.
В 1994 году этой расе охотников за инопланетянами было дано имя — Яутжа. Это имя никогда не используется в фильмах, но оно было введено в более широкую вселенную в романе «Чужой против Хищника: Добыча » и теперь широко используется фанатами. То же самое относится и к именам самих Хищников — многие из них просто использовались на съемочной площадке или были применены фанатами после выхода фильмов, но теперь это общепринятые названия.
Благодаря последнему фильму «Хищник », который выйдет в кинотеатрах 14 сентября, мы скоро сможем увидеть, какой будет следующая стадия эволюции «Хищника». Мы уже знаем из трейлеров, что режиссер Шейн Блэк приготовил что-то очень большое и страшное. Итак, давайте оглянемся на прошлый 31 год эволюции Хищника, чтобы увидеть, как именно изменился этот столь любимый (и страшный) монстр из кино. Давайте охотиться!
Ознакомьтесь с лучшими предложениями по коммутаторам, доступными прямо сейчас
Лучшие предложения для PS5, доступные прямо сейчас: экономьте на играх, аксессуарах и не только
Этот Хищник охотился и убивал почти весь военный отряд в джунглях Южной Америки. Охотник джунглей остается самым культовым из всех Хищников, начиная с биомаски, плазмозаклинателя на плече и саморазрушающейся перчатки на запястье и заканчивая ружьем и наручными клинками. Изначально шлем был более сложным, но продюсер Джоэл Сильвер подумал, что он уменьшит воздействие, когда Хищник покажет свое истинное лицо, поэтому его упростили. Нет никаких сомнений в том, что чистая эффективность конструкции является частью того, что сделало Predator поистине классическим монстром.
Нет никаких сомнений в том, что чистая эффективность конструкции является частью того, что сделало Predator поистине классическим монстром.
2. Городской Охотник
Хищник 2 (1990)
Хищник 2 показал, что существует гораздо больше видов Хищника, и начал постепенную эволюцию его внешнего вида как под броней, так и над ней. В фильме рассказывается о группе новых охотников, иногда известных как «Затерянное племя». Они прибыли в Лос-Анджелес в 1997 году и занялись своим делом. Городской охотник является главным охотником и имеет более полный набор оружия, чем его коллега из джунглей, включая сетевое ружье, комбистик (выдвижную боевую палку) и смарт-диск, культовый метательный диск Хищника. Различные члены этой охотничьей группы показали, что не все Хищники выглядят одинаково, и у гримера Стэна Уинстона было больше времени на разработку дизайна существ, чем в первом фильме. Биомаска Городского Охотника более угловатая, цвет его кожи темнее, а на лбу выделяется характерный рогатый гребень.
3. Greyback/Elder Predator
Predator 2 (1990)
Охотников Predator 2 возглавляет Greyback. Этот пожилой воин — Старейшина, который в мифологии Хищников — ветеран Яутжа, завоевавший уважение убийствами и трофеями, но больше не принимающий участия в бою. У Грейбека более длинные дреды и сломанный бивень; Интересно, что костюм, который носят в фильме, на самом деле был модифицированной версией одного из костюмов Хищника, использованного в первом фильме.
4. Dachande
Alien vs. Predator #0-4 (1990)
Один из самых известных Хищников в более широкой вселенной, Dachande (что означает «другой нож») появился в оригинальном Alien vs. Predator Комикс . Этот Яутжа, также известный как Сломанный Бивень из-за повреждения лица, является умным и благородным воином, ответственным за множество убийств ксеноморфов. Хищники из комиксов AvP продемонстрировали несколько отличий в доспехах и вооружении. Дачанде была одета в тяжелую броню, украшенную черепом ксеноморфа. В частности, эти Хищники не используют наплечный плазменный заклинатель из фильмов, вместо этого вооружены ручными бластерами.
Дачанде была одета в тяжелую броню, украшенную черепом ксеноморфа. В частности, эти Хищники не используют наплечный плазменный заклинатель из фильмов, вместо этого вооружены ручными бластерами.
5. Плохая кровь Хищник
Хищник: Плохая кровь #1-4 (1993)
Хотя Хищники безжалостны и смертоносны, тем не менее, они связаны строгим кодексом чести. Плохая кровь Хищник — сумасшедший мошенник Яутжа, который освободился от кода и начал серию убийств на улицах Нью-Джерси в комиксе Плохая кровь , в конечном итоге унеся более 50 жертв, прежде чем он был остановлен Enforcer Predator. Внешний вид Дурной Крови отмечен тем, как он украшает свои доспехи «трофейными» частями своей добычи, в том числе отрубленными головами, свисающими с его пояса, и доспехами, сделанными из костей (включая череп Хищника).
6. Young Blood Predator
Alien vs. Predator (2004)
Young Blood — Хищник-подросток, недавно прошедший обучение и готовый к Охоте. В Чужой против Хищника Молодой Блад, известный как Шрам, возглавляет двух других — Селтика и Чоппера — с миссией убить Ксеноморфа и получить статус полнокровного. Стэн Уинстон не вернулся для третьего фильма, поэтому студия VFX Amalgamated Dynamics взяла на себя продолжение эволюции Хищника. Биомаска Шрама напоминает оригинального Охотника в джунглях, но он вооружен новым оружием, в том числе более гладким комбистиком, метательными звездами и телескопическими клинками на запястьях, длина которых вдвое превышает нормальную. Шрам также является единственным Хищником, которого мы видим без маски в 9.0009 AvP , и ему была придана более человеческая окраска кожи, чтобы сделать его более симпатичным персонажем, чем в предыдущих версиях.
В Чужой против Хищника Молодой Блад, известный как Шрам, возглавляет двух других — Селтика и Чоппера — с миссией убить Ксеноморфа и получить статус полнокровного. Стэн Уинстон не вернулся для третьего фильма, поэтому студия VFX Amalgamated Dynamics взяла на себя продолжение эволюции Хищника. Биомаска Шрама напоминает оригинального Охотника в джунглях, но он вооружен новым оружием, в том числе более гладким комбистиком, метательными звездами и телескопическими клинками на запястьях, длина которых вдвое превышает нормальную. Шрам также является единственным Хищником, которого мы видим без маски в 9.0009 AvP , и ему была придана более человеческая окраска кожи, чтобы сделать его более симпатичным персонажем, чем в предыдущих версиях.
7. Ancient Predator
Alien vs. Predator (2004)
Древний Хищник даже старше Старейшины, и его можно увидеть в конце Alien vs. Селтик и Чоппер на Землю. Он отмечен длинными серыми дредами и более волосатым лицом, чем у других Хищников.
8. Волк/Элитный Хищник
Чужой против Хищника: Реквием (2007)
Волк — главный Хищник в Чужой против Хищника: Реквием . АвП. Элитный Хищник — это хорошо обученный Яутжа, а высокое и стройное телосложение Волка напоминает охотника из джунглей из первого фильма. Шлем вдохновлен городским охотником Predator 2 , но с добавлением ряда шипов, а его лицо более плоское, с острыми зубами и заметным кислотным ожогом с одной стороны (предположительно, из-за встречи с Ксеноморфом). Что касается вооружения, то Вольф пополнил арсенал Хищника некоторыми новшествами. Есть Power Gauntlet, который дает мощную ударную способность, лазерный луч, проецирующий мины, растворяющую кислоту и хлыст.
9. Супер Хищник
Хищники (2010)
Хищник продолжал развиваться, и в 2010 году был представлен новый тип Хищников . Хотя эти грозные воины того же роста, что и Хищник джунглей, они сильнее и мускулистее, с чешуйчатой кожей рептилий, зачесанными назад дредами и более длинным, похожим на морду лицом. В фильме есть три суперхищника, каждый с улучшенным вооружением и техникой охоты. Следопыт использует устрашающих охотничьих собак, известных как адские гончие, в то время как у Сокольника есть (да!) механический сокол, чтобы выслеживать свою добычу. Самым страшным из всех является Бесеркер — к его шлему добавлена тревожная костяная челюсть, похожая на череп, его биомаска позволяет ему определить пульс своей добычи, а его плазменный заклинатель — скорострельное вращающееся оружие смерти.
В фильме есть три суперхищника, каждый с улучшенным вооружением и техникой охоты. Следопыт использует устрашающих охотничьих собак, известных как адские гончие, в то время как у Сокольника есть (да!) механический сокол, чтобы выслеживать свою добычу. Самым страшным из всех является Бесеркер — к его шлему добавлена тревожная костяная челюсть, похожая на череп, его биомаска позволяет ему определить пульс своей добычи, а его плазменный заклинатель — скорострельное вращающееся оружие смерти.
10. Lord
Видеоигра Alien vs. Predator (2010)
Дополнение Sega 2010 года к серии игр AvP (также известное как AVP3 ) было не очень хорошим, но оно представило классный новый стиль био- маску одному из своих Хищников. Лорд был первым Яутжа, убившим Ксеноморфа, и в отличие от других последующих Хищников, чьи шлемы полностью закрывали рот, у этого было отверстие, через которое были видны его челюсти.
11. Хищник-мутант
Чужие против Хищника: Огонь и Камень #1-4 (2014)
Даже по меркам Хищников этот грозный мутант — страшный противник. Он фигурирует в комиксе Aliens vs. Predator: Fire & Stone и является старейшиной, подвергшейся воздействию химического вещества, ускоряющего генетику. Хищник-Мутант возвышается над другими Яутжа, обладая невероятной силой и массой челюстей, способных проламывать черепа его врагов.
Он фигурирует в комиксе Aliens vs. Predator: Fire & Stone и является старейшиной, подвергшейся воздействию химического вещества, ускоряющего генетику. Хищник-Мутант возвышается над другими Яутжа, обладая невероятной силой и массой челюстей, способных проламывать черепа его врагов.
12. Гигантский гибридный хищник
Хищник (2018)
С последним фильмом в серии, Хищник , до релиза еще пара недель, мы не совсем уверены, каким будет следующий этап эволюции Хищника. Тем не менее, трейлеры, выпущенные до сих пор, предполагают, что мы собираемся получить самый большой; существо, которое стоит на высоте 10 футов и без проблем разбрасывает меньших Хищников. Официальный синопсис фильма дает нам представление о том, чего ожидать: «Теперь самые смертоносные охотники во Вселенной стали сильнее, умнее и смертоноснее, чем когда-либо прежде, поскольку они генетически обновили себя ДНК других видов».
Хищники модифицируют эволюционную реакцию добычи на изменение температуры
Biol Lett. 2015 декабрь; 11(12): 20150798.
2015 декабрь; 11(12): 20150798.
doi: 10.1098/rsbl.2015.0798
Информация об авторе Примечания к статье Информация об авторских правах и лицензии Отказ от ответственности
- Дополнительные материалы
Поскольку климатические режимы меняются во многих экосистемах по всему миру, эволюция может стать критическим процессом, обеспечивающим устойчивость в быстро меняющихся условиях. Организмы регулярно взаимодействуют с другими видами, однако не совсем понятно, может ли эволюция, опосредованная климатом, происходить в контексте взаимодействия видов. Мы проверили, может ли взаимодействие видов изменить эволюционную реакцию на температуру. Мы демонстрируем, что давление хищников со стороны личинок двукрылых ( Chaoborus americanus ) модифицировал эволюционную реакцию пресноводных ракообразных ( Daphnia pulex ) на тепловую среду на протяжении примерно семи поколений в лабораторных условиях. Daphnia , содержащиеся при 21°C, развивались с более высокими темпами роста популяции, чем те, которые содержались при 18°C, но только в тех популяциях, которые также выращивались вместе с хищниками. Кроме того, отбор, опосредованный хищниками, привел к эволюции повышенной термической пластичности Daphnia . Этот лабораторный эксперимент по естественному отбору демонстрирует, что биотические взаимодействия могут изменять эволюционную адаптацию к температуре. Понимание взаимодействия между многочисленными селективными силами может улучшить прогнозы экологических и эволюционных реакций организмов на быстрые изменения окружающей среды.
Daphnia , содержащиеся при 21°C, развивались с более высокими темпами роста популяции, чем те, которые содержались при 18°C, но только в тех популяциях, которые также выращивались вместе с хищниками. Кроме того, отбор, опосредованный хищниками, привел к эволюции повышенной термической пластичности Daphnia . Этот лабораторный эксперимент по естественному отбору демонстрирует, что биотические взаимодействия могут изменять эволюционную адаптацию к температуре. Понимание взаимодействия между многочисленными селективными силами может улучшить прогнозы экологических и эволюционных реакций организмов на быстрые изменения окружающей среды.
Ключевые слова: адаптация, экология сообщества, дафния , эволюция, хищник, температура к изменяющейся окружающей среде может столкнуться с вызванным климатом вымиранием популяции [2]. Быстрая адаптация к переменным, связанным с климатом, была задокументирована в нескольких одновидовых системах [3–7], но, поскольку практически все организмы испытывают регулярные межвидовые взаимодействия [8], понимание того, как биотические взаимодействия влияют на эволюционные реакции, имеет решающее значение.
Контекст сообщества может влиять на эволюционные реакции, изменяя доступность генетической изменчивости или силу естественного отбора [8,9]. Кроме того, биотические взаимодействия могут ослабить эволюционные реакции на обусловленный климатом отбор, если взаимодействующие виды отбирают организмы, хорошо приспособленные к абиотической среде [10]. И наоборот, биотические взаимодействия могут усилить эволюционный ответ на климато-опосредованный отбор, если и биотическое давление, и климатическая переменная благоприятствуют сходным признакам [11]. Учитывая повсеместное распространение биотических взаимодействий в природе, наша способность предсказывать, могут ли популяции адаптироваться к быстрым изменениям окружающей среды, зависит от лучшего понимания того, ускоряют ли и каким образом биотические процессы эволюционную адаптацию к переменным, связанным с климатом [2,8,10]. ].
Мы проверили гипотезу о том, что биотические взаимодействия (хищничество) могут изменять эволюционные реакции на абиотические факторы (температуру), используя лабораторный эксперимент по естественному отбору. Личинки Chaoborus americanus (далее Chaoborus ) являются хищниками Daphnia pulex (далее Daphnia ) и, как было показано, оказывают сильный отбор на Daphnia [12]. Мы ожидали, что дафний , выращенных вместе с хищниками, будут иметь меньшие размеры тела и более высокие темпы роста популяции [13,14]. Точно так же мы ожидали Daphnia выращивалась при более высоких температурах, чтобы развить меньшие размеры тела и более высокие темпы роста [15,16]. Поскольку отбор хищников, по-видимому, влияет на жизненный цикл жертвы [13], тогда как отбор по температуре, по-видимому, влияет на размер клетки и генома [17], мы предположили, что эти два механизма отбора были относительно независимыми, и, таким образом, в присутствии обоих силы отбора, общая эволюционная реакция будет больше, чем реакция только на температуру или хищничество.
Личинки Chaoborus americanus (далее Chaoborus ) являются хищниками Daphnia pulex (далее Daphnia ) и, как было показано, оказывают сильный отбор на Daphnia [12]. Мы ожидали, что дафний , выращенных вместе с хищниками, будут иметь меньшие размеры тела и более высокие темпы роста популяции [13,14]. Точно так же мы ожидали Daphnia выращивалась при более высоких температурах, чтобы развить меньшие размеры тела и более высокие темпы роста [15,16]. Поскольку отбор хищников, по-видимому, влияет на жизненный цикл жертвы [13], тогда как отбор по температуре, по-видимому, влияет на размер клетки и генома [17], мы предположили, что эти два механизма отбора были относительно независимыми, и, таким образом, в присутствии обоих силы отбора, общая эволюционная реакция будет больше, чем реакция только на температуру или хищничество.
Мы провели полностью факторный лабораторный эксперимент по естественному отбору с тремя температурными (18°, 21° и 25°C) и двумя обработками хищниками (с одной личинкой Chaoborus или без нее). Подробная информация об обработке, а также об источнике и содержании организмов доступна в электронном дополнительном материале. Мы создали 24 микрокосма объемом 1 л (по четыре на процедуру), каждый из которых содержал 30 Daphnia и 900 мл отфильтрованной прудовой воды. Эта плотность аналогична используемой в [12]. Daphnia потребляла популяции водорослей, завезенных с водой пруда. Хотя мы не проводили количественную оценку концентрации водорослей, мы ожидали, что размер популяции Daphnia будет колебаться в зависимости от наличия пищи, как это наблюдается в природе. Питательные вещества добавлялись в микрокосмы каждые две недели в виде гранул медленного высвобождения (Osmocote ® Корм для растений в помещении и на открытом воздухе, 19-6-12). Микрокосмы испытали искусственное освещение полного спектра с циклом 14 L: 10 D.
Подробная информация об обработке, а также об источнике и содержании организмов доступна в электронном дополнительном материале. Мы создали 24 микрокосма объемом 1 л (по четыре на процедуру), каждый из которых содержал 30 Daphnia и 900 мл отфильтрованной прудовой воды. Эта плотность аналогична используемой в [12]. Daphnia потребляла популяции водорослей, завезенных с водой пруда. Хотя мы не проводили количественную оценку концентрации водорослей, мы ожидали, что размер популяции Daphnia будет колебаться в зависимости от наличия пищи, как это наблюдается в природе. Питательные вещества добавлялись в микрокосмы каждые две недели в виде гранул медленного высвобождения (Osmocote ® Корм для растений в помещении и на открытом воздухе, 19-6-12). Микрокосмы испытали искусственное освещение полного спектра с циклом 14 L: 10 D.
В конце 14-й недели (7,5 ± 1,5 Daphnia генераций [12]), хищников удаляли, а все микрокосмы помещали в общую среду (около 20°C) на три недели (недели 15–17, примерно 2–3 поколения) для минимизации материнских эффектов [12]. ]. Мы ожидали незначительных сигналов хищников в обычной садовой среде [18]. Оценка признаков приспособленности Daphnia после этой фазы общей среды позволяет нам отделить генетические изменения от фенотипической пластичности [19].
]. Мы ожидали незначительных сигналов хищников в обычной садовой среде [18]. Оценка признаков приспособленности Daphnia после этой фазы общей среды позволяет нам отделить генетические изменения от фенотипической пластичности [19].
Мы проверили, произошли ли эволюционные изменения в течение 0–14 недель, проверив, были ли изменения в двух переменных отклика— 9Скорость роста популяции 0009 Daphnia между 17 и 19 неделями и размер тела Daphnia на 19 неделе в значительной степени объяснялись селекционным воздействием. Для этого на 17-й неделе каждый микрокосм был разделен на субпопуляции ( n = 4–6 субпопуляций на микрокосм в зависимости от размера популяции микрокосма) и по этим субпопуляциям оценивались эволюционные изменения. Мы использовали низкие начальные плотности субпопуляции (0,001–0,025 отдельных мл -1 ), чтобы свести к минимуму эффекты, зависящие от плотности. Субпопуляции содержали при 18°C или 21°C (температура анализа) в течение двух недель (недели 18 и 19). ). Мы не проводили анализ эволюции с хищниками из-за недостаточного размера выборки.
). Мы не проводили анализ эволюции с хищниками из-за недостаточного размера выборки.
Мы регистрировали размер популяции Daphnia каждые две недели с 0 по 14 неделю путем усреднения количества Daphnia в трех образцах по 100 мл, взятых из каждого микрокосма. В анализе эволюции мы зафиксировали размер субпопуляции Daphnia и средний размер тела (мм 2 ) в конце 19-й недели. Площадь боковой поверхности Daphnia была измерена с использованием трех особей каждой субпопуляции, сделанных сбоку (Leica). Комплект приложений версии 4.3.0). Мы рассчитали на душу населения скорость роста, r на подгруппу населения, как численность населения на 19-й неделе минус численность субпопуляции на 17-й неделе, деленная на численность субпопуляции на 17-й неделе. Мы разделили это значение на 14 дней, чтобы рассчитать количество новых особей производится на особь дафний в сутки.
Мы использовали дисперсионный анализ, чтобы выяснить, объясняют ли температура и хищники изменение среднего размера популяции Daphnia в течение недель 0–14. Мы использовали линейные модели со смешанными эффектами для оценки важности «температуры отбора», «хищничества отбора» и «температуры анализа» (все фиксированные эффекты) в прогнозировании эволюции 9.0009 Daphnia r и размер тела. Окончательная модель для обоих признаков Daphnia включала три основных эффекта: гармонический средний размер популяции, микрокосм (случайный эффект) и взаимодействия температуры отбора × хищничества отбора и хищничества отбора × температуры анализа (подробности выбора модели см. в электронном дополнительном материале). ). Все статистические анализы проводились в R [20] с использованием пакетов car и nlme [21,22].
Мы использовали линейные модели со смешанными эффектами для оценки важности «температуры отбора», «хищничества отбора» и «температуры анализа» (все фиксированные эффекты) в прогнозировании эволюции 9.0009 Daphnia r и размер тела. Окончательная модель для обоих признаков Daphnia включала три основных эффекта: гармонический средний размер популяции, микрокосм (случайный эффект) и взаимодействия температуры отбора × хищничества отбора и хищничества отбора × температуры анализа (подробности выбора модели см. в электронном дополнительном материале). ). Все статистические анализы проводились в R [20] с использованием пакетов car и nlme [21,22].
В целом, мы обнаружили, что (i) хищники и температура опосредовали 9Динамика популяции 0009 Daphnia , (ii) Daphnia эволюционировала быстрее при выращивании с хищниками и при более высокой температуре, и (iii) отбор с хищниками привел к эволюции Daphnia меньшего размера тела.
Daphnia средний размер популяции между 0 и 14 неделями был самым высоким при 21°C ( F 2,18 = 7, p = 0,006; , вставка). При всех температурах хищники сократили размер популяции Daphnia примерно на 36% (9).0009 Ф 1,18 = 6,4, р = 0,02; , вставка). Все популяции, выращенные при 25°С, вымерли к 10-й неделе эксперимента. Опосредованное хищником сокращение размера популяции добычи, вероятно, было связано с прямым хищничеством и поведением избегания хищника [23].
При всех температурах хищники сократили размер популяции Daphnia примерно на 36% (9).0009 Ф 1,18 = 6,4, р = 0,02; , вставка). Все популяции, выращенные при 25°С, вымерли к 10-й неделе эксперимента. Опосредованное хищником сокращение размера популяции добычи, вероятно, было связано с прямым хищничеством и поведением избегания хищника [23].
Открыть в отдельном окне
Влияние режима селекции на численность популяции Daphnia . Светло-синий, темно-синий и зеленый символы обозначают температурные обработки 18°, 21° и 25°C соответственно. Кружки/сплошные линии и треугольники/пунктирные линии обозначают режимы селекции в отсутствие и в присутствии хищников. Столбики погрешностей составляют ± 1 сем. Обратите внимание, что статистический анализ проводился на основе средних значений (вставка) и что недельные значения включены сюда для полноты картины. (Онлайн-версия в цвете.)
Дафнии , выращенные при 21°C, развивались с более высокими темпами роста, чем дафнии, выращенные при 18°C, но только в том случае, если они также выращивались с хищниками ( a и ). Этот результат может быть связан с повышенной скоростью атаки Chaoborus при более высоких температурах [24], что приводит к более сильному отбору для увеличения скорости роста при более высокой температуре отбора. Неясно, могла ли Daphnia , выращенная в отсутствие хищников, эволюционировать к изменению температуры, если бы мы проводили эксперимент дольше. Мы также заметили, что Дафния , отобранная вместе с хищниками, показала более быстрый рост при более высокой температуре анализа ( a и ). Это эволюционное увеличение пластичности поддерживает утверждение о том, что биотические взаимодействия могут быть важными движущими силами эволюционных изменений норм реакции [25]. Потенциальный механизм этого результата заключается в том, что обработка хищником создавала более гетерогенную среду в отношении пространственного или временного распределения риска хищничества, чем обработка температурой, а переменная среда была связана с эволюцией повышенной пластичности [26].
Этот результат может быть связан с повышенной скоростью атаки Chaoborus при более высоких температурах [24], что приводит к более сильному отбору для увеличения скорости роста при более высокой температуре отбора. Неясно, могла ли Daphnia , выращенная в отсутствие хищников, эволюционировать к изменению температуры, если бы мы проводили эксперимент дольше. Мы также заметили, что Дафния , отобранная вместе с хищниками, показала более быстрый рост при более высокой температуре анализа ( a и ). Это эволюционное увеличение пластичности поддерживает утверждение о том, что биотические взаимодействия могут быть важными движущими силами эволюционных изменений норм реакции [25]. Потенциальный механизм этого результата заключается в том, что обработка хищником создавала более гетерогенную среду в отношении пространственного или временного распределения риска хищничества, чем обработка температурой, а переменная среда была связана с эволюцией повышенной пластичности [26].
Открыть в отдельном окне
Влияние температуры селекции, хищничества селекции и температуры анализа на Daphnia ( a ) скорость роста на душу населения , r и ( b ) размер тела. Синие символы (штриховые линии) и голубые символы (сплошные линии) обозначают отбор с хищниками и без них соответственно. Кружками и треугольниками обозначены популяции, отобранные при 18°C и 21°C соответственно. Столбики погрешностей составляют ± 1 сем. (Онлайн-версия в цвете.)
Таблица 1.
Влияние режима селекции и температуры анализа на скорость роста Daphnia per capita ( r ) и размер тела. Представлены значения Wald χ 2 и p из моделей смешанных эффектов. Значения, выделенные курсивом, обозначают статистически значимые параметры.
| на душу населения скорость роста | размер тела | |||
|---|---|---|---|---|
| p | p | |||
| selection temperature | 0. 070 070 | 0.791 | 0.011 | 0.917 |
| selection predation | 8.638 | 0.003 | 3 . 943 | 0 . 047 |
| температура анализа | 2,163 | 0,141 | 3,563 | 0.059 |
| population size | 2.641 | 0.104 | 1.428 | 0.232 |
| selection predation × assay temperature | 8. 797 | 0 . 003 | 1,128 | 0,289 |
| температура выбора × предыдущая выборка | 9,755 | 0 0 002 | 0,0012 | 0.973 |
Открыть в отдельном окне
Дафнии , выращенные с хищниками, развились в меньших размерах тела, чем выращенные без хищников ( b и ). Этот результат противоречит некоторой теории жизненного цикла этой пары хищник-жертва [27], но согласуется с теорией хищничества, не зависящего от размера [14]. Эти результаты также не согласуются с аналогичным экспериментом с Chaoborus и Daphnia [12], но там Daphnia испытывала постоянную доступность пищи и в среднем была больше по размеру, чем в этом эксперименте. Будущие исследования, изучающие поведение хищников при кормлении при разных температурах, помогут выяснить, способствовало ли избирательное по размеру хищничество наблюдаемой здесь модели размеров тела. Не было влияния температуры селекции на размер тела Daphnia , и Daphnia , проанализированные при 21°C, были немного меньше, чем проанализированные при 18°C ( b и ). Эта пластичность размера тела согласуется с предыдущим исследованием, в котором изучались пластические реакции Daphnia в ответ на изменение температуры [28].
Будущие исследования, изучающие поведение хищников при кормлении при разных температурах, помогут выяснить, способствовало ли избирательное по размеру хищничество наблюдаемой здесь модели размеров тела. Не было влияния температуры селекции на размер тела Daphnia , и Daphnia , проанализированные при 21°C, были немного меньше, чем проанализированные при 18°C ( b и ). Эта пластичность размера тела согласуется с предыдущим исследованием, в котором изучались пластические реакции Daphnia в ответ на изменение температуры [28].
Этот эксперимент показал, что давление хищников опосредовало эволюционное расхождение Daphnia в различных температурных условиях. Подтверждение этих результатов в полевых условиях помогло бы оценить их универсальность. Учитывая, что многие хищники, по прогнозам, будут выбирать более быструю скорость роста добычи [14] и что повышенные температуры имеют тенденцию увеличивать скорость роста экотерм [15,16], синергетические взаимодействия между температурой и хищниками могут представлять собой важный механизм адаптации организмов к быстрому изменению климата. меняющаяся среда. Наши результаты дополняют растущий объем экологической литературы о сохранении популяций в контексте взаимодействия видов в меняющихся условиях [29].,30], демонстрируя возможные эволюционные преимущества трофических взаимодействий для быстрой адаптации в условиях потепления. Кроме того, эти результаты демонстрируют, что эволюционные изменения могут происходить в масштабах экологического времени, и, таким образом, усиливают аргументы в пользу включения эволюционных процессов в объяснения краткосрочных экологических паттернов [31].
меняющаяся среда. Наши результаты дополняют растущий объем экологической литературы о сохранении популяций в контексте взаимодействия видов в меняющихся условиях [29].,30], демонстрируя возможные эволюционные преимущества трофических взаимодействий для быстрой адаптации в условиях потепления. Кроме того, эти результаты демонстрируют, что эволюционные изменения могут происходить в масштабах экологического времени, и, таким образом, усиливают аргументы в пользу включения эволюционных процессов в объяснения краткосрочных экологических паттернов [31].
Tseng & O’Connor Дополнительные материалы-методы и рисунок S1. Фото дафнии:
Нажмите здесь для просмотра. (146K, pdf)
Мы благодарим Н. Колка за помощь в настройке и обслуживании эксперимента, а также Л. Ризеберга и Р. Стокса за полезные комментарии. Мы также благодарим Дж. Петкау, Д. Динсдейла и выпускников STAT551 Statistical Consulting в UBC за обзор этого проекта.
Исследуемые организмы были собраны в экспериментальных прудах Университета Британской Колумбии. Никаких разрешений не требовалось.
Данные доступны в цифровом репозитории дриад: http://dx.doi.org/10.5061/dryad.m64qc.
М.Т. задумал исследование. М.Т. и М.И.О. разработал эксперимент. М.Т. собрал и проанализировал данные. М.Т. написал рукопись. М.И.О. редактировал рукопись. Оба автора дали окончательное согласие на публикацию.
У нас нет конкурирующих интересов.
Финансирование было предоставлено грантом NSERC (Канада) Discovery компании M.I.O.
1. МГЭИК. 2014. Резюме для политиков. В «Изменении климата, 2014 г.: последствия, адаптация и уязвимость». Часть A: глобальные и отраслевые аспекты. Вклад Рабочей группы II в Пятый оценочный доклад Межправительственной группы экспертов по изменению климата Кембридж, Великобритания: Издательство Кембриджского университета. [Академия Google]
2. Berg MP, Kiers ET, Driessen G, van der Heijden M, Kooi BW, Kuenen F, Liefting M, Verhoef HA, Ellers J. 2010.
Адаптироваться или рассредоточиться: понимание устойчивости видов в меняющемся мире. Глоб. Изменить биол. 16, 587–598. ( 10.1111/j.1365-2486.2009.02014.x) [CrossRef] [Google Scholar]
2010.
Адаптироваться или рассредоточиться: понимание устойчивости видов в меняющемся мире. Глоб. Изменить биол. 16, 587–598. ( 10.1111/j.1365-2486.2009.02014.x) [CrossRef] [Google Scholar]
3. Huey RB, Kingsolver JG. 1993. Эволюция устойчивости к высокой температуре у экзотермов. Являюсь. Нац. 142, С21–С46. ( 10.1086/285521) [CrossRef] [Google Scholar]
4. Lenski RE, Bennett AF. 1993. Эволюционная реакция Escherichia coli на тепловой стресс. Являюсь. Нац. 142, С47–С64. ( 10.1086/285522) [PubMed] [CrossRef] [Google Scholar]
5. Партридж Л., Барри Б., Бартон Н.Х., Фаулер К., Френч В. 1995. Быстрая лабораторная эволюция признаков взрослой жизни у Drosophila melanogaster в ответ на температуру. Эволюция 49, 538–544. ( 10.2307/2410277) [PubMed] [CrossRef] [Google Scholar]
6. Van Doorslaer W, Stoks R, Duvivier C, Bednarska A, De Meester L.
2009 г..
Динамика популяции определяет генетическую адаптацию к температуре у дафний . Эволюция
63, 1867–1878 гг. ( 10.1111/j.1558-5646.2009.00679.x) [PubMed] [CrossRef] [Google Scholar]
( 10.1111/j.1558-5646.2009.00679.x) [PubMed] [CrossRef] [Google Scholar]
7. Коллинз С. 2011. Конкуренция ограничивает адаптацию и продуктивность фотосинтезирующих водорослей при повышенном уровне CO 2 . проц. Р. Соц. Б 278, 247–255. ( 10.1098/rspb.2010.1173) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
8. Johnson MTJ, Stinchcombe JR. 2007. Новый синтез между экологией сообщества и эволюционной биологией. Тенденции Экол. Эвол. 22, 250–257. ( 10.1016/j.tree.2007.01.014) [PubMed] [CrossRef] [Google Scholar]
9. Фрэнкхэм Р. 1996. Отношение генетической изменчивости к размеру популяции в дикой природе. Консерв. биол. 10, 1500–1508 гг. ( 10.1046/j.1523-1739.1996.10061500.x) [CrossRef] [Google Scholar]
10. Strauss SY. 2014. Экологические и эволюционные реакции в сложных сообществах: последствия для вторжений и экоэволюционных обратных связей. Ойкос 123, 257–266. ( 10.1111/j.1600-0706.2013.01093.x) [CrossRef] [Google Scholar]
11. Crain CM, Kroeker K, Halpern BS.
2008.
Интерактивное и кумулятивное воздействие нескольких факторов стресса человека в морских системах. Экол. лат. 11, 1304–1315. ( 10.1111/j.1461-0248.2008.01253.x) [PubMed] [CrossRef] [Google Scholar]
Crain CM, Kroeker K, Halpern BS.
2008.
Интерактивное и кумулятивное воздействие нескольких факторов стресса человека в морских системах. Экол. лат. 11, 1304–1315. ( 10.1111/j.1461-0248.2008.01253.x) [PubMed] [CrossRef] [Google Scholar]
12. Шпитце К. 1991. Хищничество Chaoborus и эволюция жизненного цикла Daphnia pulex: временная картина разнообразия популяции, приспособленности и средней продолжительности жизни. Эволюция 45, 82–92. ( 10.2307/2409484) [PubMed] [CrossRef] [Google Scholar]
13. Riessen HP. 1999. Вызванные хищниками изменения в жизненном цикле Daphnia : синтез исследований с использованием метаанализа. Может. Дж. Фиш. Аква. науч. 56, 2487–2494. ( 10.1139/cjfas-56-12-2487) [CrossRef] [Google Scholar]
14. Abrams PA, Rowe L. 1996. Влияние хищничества на возраст и размер зрелости добычи. Эволюция 50, 1052–1061. ( 10.2307/2410646) [PubMed] [CrossRef] [Google Scholar]
15. Аткинсон Д.
1994.
Температура и размер организма: биологический закон для экзотерм?
Доп. Экол. Рез. 25, 1–58. ( 10.1016/S0065-2504(08)60212-3) [CrossRef] [Google Scholar]
Экол. Рез. 25, 1–58. ( 10.1016/S0065-2504(08)60212-3) [CrossRef] [Google Scholar]
16. Angilletta MJ. 2009. Термическая адаптация: теоретический и эмпирический синтез. Оксфорд, Великобритания: Издательство Оксфордского университета. [Google Scholar]
17. Джалал М., Воеводзич М.В., Лаане К.М.М., Хессен Д.О. 2013. Больше Daphnia при более низкой температуре: роль размера клетки и генома. Геном 519, 511–519. ( 10.1139/gen-2013-0004) [PubMed] [CrossRef] [Google Scholar]
18. Додсон С. 1988 год. Экологическая роль химических раздражителей для зоопланктона: поведение избегания хищников у Daphnia . Лимнол. океаногр. 33, 1431–1439. ( 10.4319/lo.1988.33.6_part_2.1431) [CrossRef] [Google Scholar]
19. Mitchell SE, Lampert W. 2000. Температурная адаптация у географически распространенного зоопланктера, Дафния большая . Дж. Эвол. биол. 13, 371–382. ( 10.1046/j.1420-9101.2000.00193.x) [CrossRef] [Google Scholar]
20. 20. R Core Team
2014. R: язык и среда для статистических вычислений. Вена, Австрия: Фонд статистических вычислений R; (http://www.R-project.org) [Google Scholar]
R: язык и среда для статистических вычислений. Вена, Австрия: Фонд статистических вычислений R; (http://www.R-project.org) [Google Scholar]
21. Fox J, Weisberg S. 2011. Компаньон {R} для прикладной регрессии. Таузенд-Оукс, Калифорния: Sage Publications. [Google Scholar]
22. Пинейро Дж., Бейтс Д., Деброй С., Саркар Д., Team RC. 2015. nlme: линейные и нелинейные модели смешанных эффектов. Пакет R версии 3.1 (http://CRAN.R-project.org/package=nlme)
23. Лагергрен Р., Леберфингер К., Стенсон Дж. Э. 2008. Сезонные и онтогенетические вариации суточной вертикальной миграции Chaoborus flavicans и их влияние на поведение другого зоопланктона при выборе глубины. Лимнол. океаногр. 53, 1083–1092. ( 10.4319/lo.2008.53.3.1083) [CrossRef] [Google Scholar]
24. Шпитце К. 1985. Функциональная реакция хищника из засады: хищничество Chaoborus americanus на Daphnia pulex . Экология 66, 938–949. ( 10.2307/1940556) [CrossRef] [Google Scholar]
25.